Team:Calgary/Part Collection

OVERVIEW
Promoting a New Sustainable Chassis: A Yarrowia lipolytica collection
To expand the accessibility and usability of synthetic biology, new methods need to be introduced to promote the sustainable growth of biological organisms. Cellulose is the most abundant organic compound on the planet (Brigham, 2018); therefore, engineering organisms to be able to grow on cellulose would be very advantageous over using expensive media. Y. lipolytica is a food-safe “non-conventional” yeast that is an up-and-coming chassis in synthetic biology, due to its ability to produce beneficial metabolites and fats (Goncalves et al, 2014).
Many cellulases derived from bacteria and fungi have been deposited to the iGEM registry over the years. However our part collection is the first of its kind in providing cellulases optimized for function in Y. lipolytica. Furthermore, we are introducing amino acid synthesis genes to create auxotrophic and overproducing strains to expand Y. lipolytica's versatility in molecular biology. We are also introducing reporter devices and an antibiotic resistance cassette to increase the usability of this chassis.
Our novel Y. lipolytica collection includes all of our parts which, in tandem with our Y. lipolytica protocols, allow Y. lipolytica to be a more accessible and usable chassis in the iGEM community and beyond.
COLLECTION COMPONENTS
Allowing for thoughtful and modular assembly
Our collection contains 26 parts, including basic parts that can be used by anyone working with Y. lipolytica, and fully functional protein generator devices containing the cellulase expression cassettes, an antibiotic resistance cassette, and amino acid synthesis cassettes. Our devices also include the gibson homology sequences for the flexible assembly. Overall, we are contributing:
- 1 Y. lipolytica signal peptide
- 2 constitutive Y. lipolytica promoters
- 1 Y. lipolytica terminator
- 4 protein coding sequences for fungal cellulases which have been codon-optimized for Y. lipolytica
- 2 novel protein coding sequences for fungal cellulases that have been modelled in-silico and modified at certain residues for better pH and temperature stability
- 6 protein generator devices containing the different cellulases with gibson homology sequences for modular assembly
- The protein coding sequence for nourseothricin resistance
- A nourseothricin resistance protein generator device
- 2 coding sequences for Y. lipolytica amino acid synthesis genes
- 2 amino acid over expression constructs
- 2 reporter genes codon optimized for Y. lipolytica
- 2 high expression reporter gene constructs
To learn more about each of the parts, check out their registry pages below.
<groupparts>iGEM20 Calgary</groupparts>Note: All devices are fully functional expression constructs that can be directly cloned and transformed.
PART DESIGN
Part features
Our parts have been designed for high protein expression and secretion (for the cellulases) in Y. lipolytica, and can even be assembled together to create larger cassettes. The cellulases can be easily purified using the 6X His tag, FLAG tag, or Myc tag (all proteins have one of these tags is present at the N-terminus) if necessary, or the tags can be used for western blot detection. A thrombin cleavage site has also been added between the tags and the coding sequence so the tag can be removed if it interferes with protein function.
Additionally, our Trichoderma reesei EGI and Penicillium funiculosum CBHI have been modified at certain residues based on our modelling for higher efficiency at the pH and temperature optimal for Y. lipolytica growth. You can read more about their design and characterization on the characterization and modelling pages.
Modular Gibson Assembly
Y. lipolytica is known to favour linear DNA integration when attempting to modify its genome. However, transforming a linear piece of DNA presents a challenge as there still needs to be a selection marker present; something that would normally be inherent to a plasmid. Therefore, parts must be attached to a selection marker to easily screen for successful transformants. Flanking restriction sites (NotI) can then be used to linearize the DNA for transformation into Y. lipolytica (figure 1).

Figure 1. Process of integrating a gene of interest in the Yarrowia lipolytica genome as a larger cassette containing an antibiotic resistance gene to select for successful transformants.
NOURSEOTHRICIN RESISTANCE CASSETTE: A KEY PART
For the selection marker to be included in the linearized DNA being transformed, multiple transcriptional units (i.e. the selection marker and the GOI) need to be assembled into one construct. After consultation with Dr. Raymond Turner and PhD student Trevor Randall, we decided to design our parts for Gibson Assembly. Since the selection marker is the common necessity for all parts, we decided to make it a destination vector. Our Nourseothricin Resistance Cassette (BBa_K3629015), when cloned into a plasmid, acts as a destination vector for multiple transcriptional units (i.e devices) to be assembled into. Digestion with SmaI exposes the ends of the Gibson 1 and Gibson 2 homology sequences for the 5' ends to be chewed back by T5 exonuclease, resulting in ~40bp ssDNA regions for incoming parts (with Gibson 1 and 2) to complement/bind to. Once the parts are added and the plasmid is recircularized, NotI can be used to linearize the DNA for transformation into Y. lipolytica (figure 2). We chose nourseothricin resistance as a selection marker by recommendation from Y. lipolytica expert Dr. Rodrigo Ledesma-Amaro , as this antibiotic is relatively cheap, the gene size is small making it easier to order/synthesize, and not many well-characterized auxotrophic Y. lipolytica strains are available yet.
All of our devices include the Gibson 1 and 2 homology sequences. These sequences have been made to have minimal secondary structures when in their ssDNA form after T5 exonuclease digestion. The Nourseothricin cassette is therefore a key component of our collection and can be used as a destination vector for all future teams working with Y. lipolytica parts as long as they have the correct Gibson homology sequences (figure 2).
- Gibson 1: 5' gggttacaggttacaatacaccaattactaatgaagtccc 3'
- Gibson 2: 5' gggtcgtttacctaactatactaaatcaActgcggttccc 3'
Figure 2. Gibson Assembly where large gene cassettes with multiple transcriptional units are assembled in a "destination vector" containing an antibiotic selection marker. Gibson 1 and Gibson 2 refer to unique homology sequences required for Gibson Assembly.
Since Y. lipolytica primarily accepts foreign DNA through non-homologous end joining, the Nourseothricin Resistance Cassette also contains flanking gPCR primer sites to sequence the surrounding area of the integration site to determine where the DNA inserted in the genome (figure 3).
- Upstream: 5' GCAGAGACTTAACATTAGCAAGC 3'
- Downstream: 5' GGTGATTGACTGATGTGAATCC 3'
We recognize however, that specifically-targeted gene integration may be necessary in some cases, and through our discussion with Dr. Rodrigo Ledesma-Amaro we learned that homologous recombination is possible with long (≥1kb) homology regions flanking the insert, which can be further enhanced through the use of Zeta sequencings. Therefore, our Nourseothricin cassette also contains another 2 sets of Gibson homology sequences (named Gibson A/B and Gibson C/D) that can be used (after digestion with HpaI) to add homology sequences after (or before) a fully assembled plasmid is made (figure 3).

Figure 3. Top: Features of Nourseothricin Resistance Cassette (BBa_K3629015) as a destination vector. Bottom: Final assembled plasmid with two GOIs in the destination vector. Gibson A/B and C/D can be exposed to T5 exonuclease activity after digestion with HpaI if genomic homology regions need to be added. gPCR primer binding sites are also highlighted.
CREATING MULTIPLE DIFFERENT PLASMIDS WITH THE SAME PARTS
For our project, we need multiple genes introduced to the Y. lipolytica genome as a single gene cassette, therefore we wanted to make our parts as modular as possible to allow for different ways of assembly for troubleshooting purposes. Multiple different Gibson homology sequences were built into the parts to allow for the assembly of different plasmids (figure 4). The ends of each Gibson homology sequence can be exposed through the use of specific restriction enzymes (see Appendix) to allow the T5 exonuclease to degrade one of the strands to create the ssDNA homology regions. Check out the registry pages for each of our expression constructs (BBa_K3629012-BBa_K3629018, BBa_K3629023-BBa_K3629026) to see which Gibson sites are built in each construct. Overall, this design allows for easier troubleshooting and provides alternate routes to success when assembling gene cassettes containing multiple transcriptional units.
Figure 4. Modular Gibson Assembly through the use of different Gibson homology sequences. Unique restriction enzymes are places at the end of each Gibson homology sequence to expose the 5' end of T5 exonuclease. This creates unique ssDNA regions that can be used to create different kinds of plasmids with the same parts.
The parts that have been designed for this kind of modular Gibson Assembly are:<.p>
- BBa_K3629013: Modified P. funiculosum CBHI expression construct
- BBa_K3629014: N. crassa CBHI expression construct
- BBa_K3629015: T. reesei CBHII expression construct
- BBa_K3629016: Modified T. reesei EGI expression construct
- BBa_K3629017: T. reesei EGII expression construct
Furthermore, the following parts have been designed to be assembled with just the Nourseothricin resistance expression construct (BBa_K3629012):
- BBa_K3629018: N. patriciarum BGS expression construct
- BBa_K3629023: 2-isopropylmalate synthase (LEU4) overexpression construct
- BBa_K3629024: Anthranilate synthase component 1 (TRP2) overexpression construct
- BBa_K3629025: mCherry expression construct
- BBa_K3629026: mCitrine expression construct
For troubleshooting purposes, the parts were also designed to be sequentially assembled if a single reaction is not working; therefore sequential integration of multiple parts may be beneficial. Our parts can be assembled in a single reaction or in sequential reactions (figure 5). Check out the appendixto see all the different possible plasmids that can be assembled using our collection.
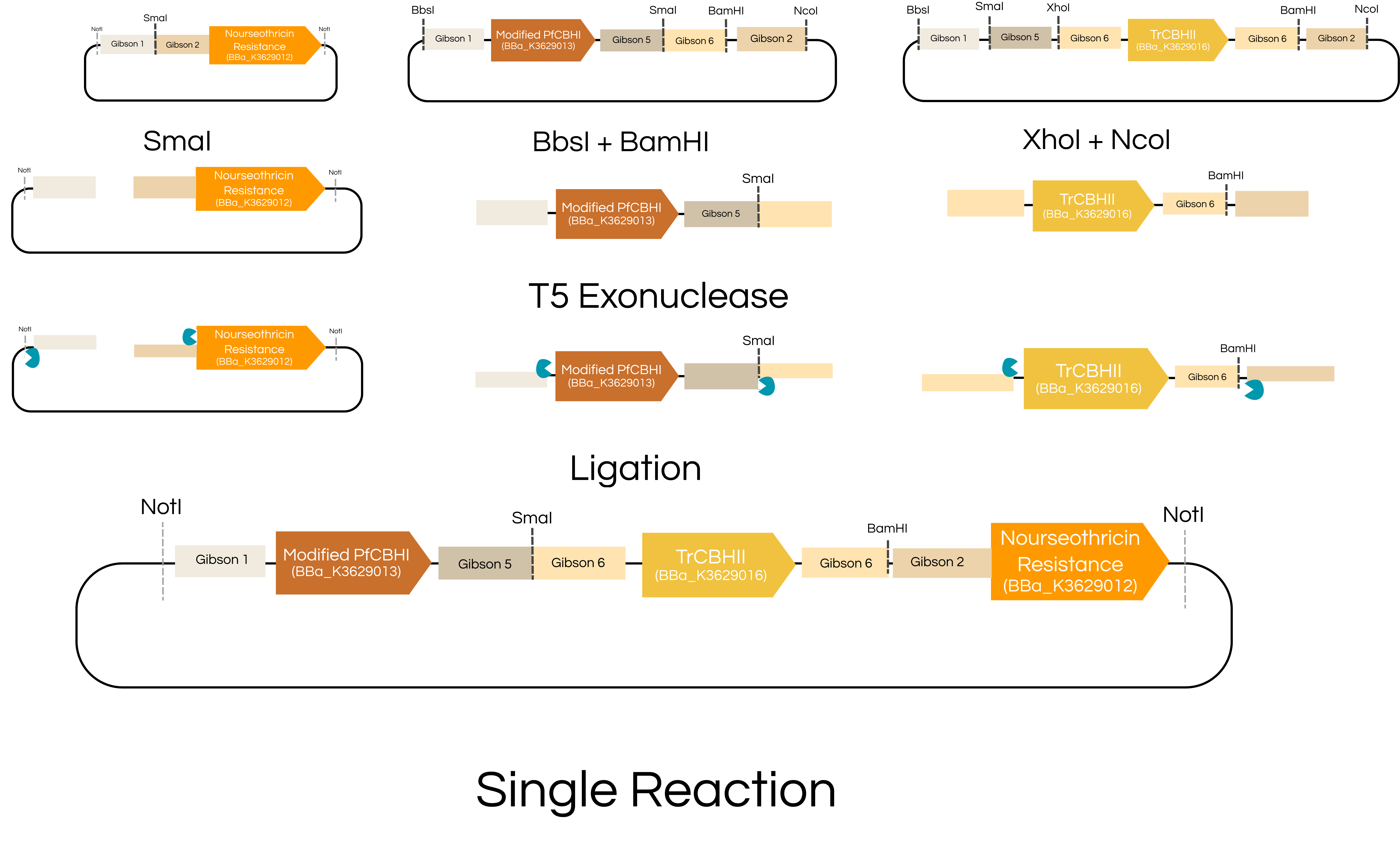

Figure 5. Gibson Assembly of multiple transcriptional units using a single reaction and in sequential reactions. Gibson 1,2, 5, and 6 refer to unique 40bp homology sequences. Click on the images to see the enlarged view (Left image= single reaction, Right image=sequential reactions).
LIST OF BUILT-IN GIBSON HOMOLOGY SEQUENCES
There are 14 different Gibson homology sequences that were used across our parts (BBa_K3629012-BBa_K3629018). Each sequence was design to have minimal secondary structure and optimal GC richness (40-60%).
Gibson 1= 5’ GGGTTACAGGTTACAATACACCAATTACTAATGAAGTCCC 3’
Gibson 2= 5’ GGGTCGTTTACCTAACTATACTAAATCAACTGCGGTTCCC 3’
Gibson 3= 5’ CCCGTAATGTGATGTCCTGTATGTTAATGCTATCGTTGGG 3’
Gibson 4= 5’ ATGAAGCTTTCCACCATCCTTTTCACAGCCTGCGCTACCCTG 3’
Gibson 5= 5’ GGGCATTACACTACAGGACATACAATTACGATAGCAACCC 3’
Gibson 6= 5’ GGGATAAGCCTATATCAGTACCTAAGCCAATCGTAACCAG 3’
Gibson 7= 5’ GGATAAATGATCCGTATGACTCATATCTTCAGCCACTGAT 3’
Gibson 8= 5’ ATCAAATCGTAATGCTTATGCCTAGTCGATCACCAGAGAT 3’
Gibson X= 5’ TTCCACCATCCTTTTCACAGCCTGCGCTACCCTG 3’
Gibson Y= 5’ GGGTTACAGGTTACAATACACCAATTACTAATGAAGTCCCGCTAG 3’
Gibson A= 5’ GGATATTACTAGGCATACTGAGTATAGAAGTCGGTGGGTT 3’
Gibson B= 5’ AACTTTAGCATTACGAATACGGATCAGCTAGACAACCTTC 3’
Gibson C= 5' CAGGATACGGATAACTACCTACACAACTAGACCTCTCGTT 3'
Gibson D= 5' AACTTCAGCGGACTAGACAAATACCATCACTGGCATAGAA 3'
The Gibson sequences present in each expression construct, and the restriction enzymes required to expose each sequence to T5 exonuclease activity are outlined on each part’s registry page. However, here is a list of the general function of each Gibson homology sequence:
- Gibson 1-2= To attach to the nourseothricin destination vector (BBa_K3629015)
- Gibson 3,5-8= Used to assemble with the different expression constructs
- Gibson X-Y= Used to switch the TEFin promoter (BBa_K3629001) with the TEF1 promoter with the partial Lip2 signal peptide (BBa_K3629003) promoter in an expression construct.
- Gibson 4= Used to attached the TEF1 promoter with a partial Lip2 signal peptide (BBa_K3629003) to a protein (coding sequence) being secreted with the Lip2 tag
- Gibson A-D= Digested with HpaI to expose ends to T5 exonuclease. Is used to add genomic homology sequences for directed insert integration.
REFERENCES
C. B., &; Dransfield, T. (2018). Chapter 3.22 - Biopolymers: Biodegradable Alternatives to Traditional Plastics, In: Green Chemistry. In 1013380455 779746459 B. Török (Ed.), Green Chemistry (Vol. 2018, pp. 753-770). Elsevier. doi:https://doi.org/10.1016/B978-0-12-809270-5.00027-3.
Gocalves, F., Colen, G., & Takahashi, J. A. (2014). Yarrowia lipolytica and Its Multiple Applications in the Biotechnological IndustryF. The Scientific World Journal, 2014. doi:https://doi.org/10.1155/2014/476207



